|
|
|
|
|
|
|
�ĥ|���G�o�f�ΧK�̾�
²��
�b�g���ʪ��y��f(epidemics)�M���@�ɪ��j�y��f(pandemics)���z�o���A
�y��ʷP�_�f�r�W�S����O�w���ҩP���a�b�����~�ֲէO���A �c�W�L���a�ް_��ʵo���I�l��(acute
febrile respiratory)�e�f�C�y��ʷP�_����ӯS�ʻ����F�\�h�f�r�b�y��f��(epidemiological)�W�������C�����O����O�b�����νު��x�éҤ��X�{�M�Ǽ��C�o��O�O�Ѱ�]����(genetic
reassortment)�Ϊ����Ǽ��Ϋ�b���W�h�����j���Ǽ���H���C�䦸�O���f�r�w�b�H���T�߫��A
�����t�M���i�w�Ƥέ��n�K�̥ؼЪ��ܭ�(antigenic)���ܡC
�@�ӯ�ް_�j�q�o�f�v(morbidity)�A ���j���w�̦��`�v�ΰ���IJ�ǬV�ʪ��f�r�O�@�ӭ쫬���J�{(archetypal
anxiety)�C�y��ʷP�_����O�h���ͳo�˪��@�ӧ����C�̦��ۤQ���@���A �����ް_�H���e�f�_�]���y��ʷP�_�f�r�w�g�Ǽ��b�H���ڸs����(Cox
& Kawaoka 1998)�A �ɭP�C�@�ܤT�~���g���y��ʵo���I�l���e�f�C���~�A �]���@�ӷs�f�r�b�@�ӨS���K�̤O���`�ڸs���X�{�A
�ҥH�C�@�ӥ@�����g����@�ǥ���y��ίਲ਼�t�a���ܥ��@�ɪ��e�f�C����y��e�f���S�x�]�A�q�`�b�u�`���~�X�{�A
���ר��t���ǬV�æb�a�y�����P�a��P���z�o�A �M�b�Ҧ��~�ֲդ������������v�A �Ʀܳs���d�~�����H���������`�v�C�Ҽ{��@�ɤH�f�A
��ڮȦ�M�ȹC�~���W����A����ʬy��ʷP�_�G���z�o�Ʀܱo��@�ӧt�Ǽ�����O�C���F������a�A�ѳo�ӥ��y�y��ʫ¯٪��I���A
�o�ӳ��`�N�|�y�z�e�f���o�f(pathogenesis)��z�M�f�r�P�K�̨t�Τ������v�ɡC
�o�f
�y��ʷP�_�f�r���P�f��(pathogenicity)�M�r�ʬO�ѴX�Ӥ��ۼv�T���]���ҨM�w:
��) �J�D�]��:
- �ؼзP�����b�J�D�ӭM�W���s�b
- �ï��b�J�D�ӭM�̪��i�ΩʬO�b�f�r���i�J�M�ƻs�����i�ʪ�
- �ӧO�J�D�K�̤O(immunocompetence)�����A
- �S���ʧK��(Specific immunity)�b�ӧO�J�D�Υؼбڸs����I�Y�f�r������Ѫ��ϰ�(epitopes)
- �K�̨t�Υѵo���Ϧ��Ħa����f�r�ƻs�ӨS����J�D�y���Y�����a�l�˪���O
�A) �f�r�]��:
- ��J�D�ӭM�����X��O
- �f�r��������O (virus shedding)
- �ӭM�f�z�ܤ�(cytopathogenic effects)������e�\�f�r�ƻs�M�ѱJ�D������A
���@�ӦX�A������
- �K�̤���������ܩ����O�X�ʧܭ��ܲ����t�ܡA �ϯ�ѧK�̺ʱ��t��(immunosurveillance)���k��
- �q�H�b�@�ͶǬV�f(zoonotic disease)�ӨӤ��P���f�r�~��(strains)�P���X�A
�ϥѧK�̺ʱ��t�Τ��k��
- �K�̤������վ��z(attenuate)���Ī��J�D���m����
�f�r���I�J: �f�r���i�J�J�D?
�y��ʷP�_�Q�ǰe���D�n�覡�O�z�L�B��(aerosols) �M�p�w(droplets)�A
�q�H�Ǩ�H�C�y��ʷP�_�M��q�L�I�l�D�i�J�J�D�C�@�ӤH���ͳ����j���T���Ӳݪw�n(terminal
sacs)�A �s���ͪw(alveoli)�A�b�Q�l���Ů�M��G����������洫���_��@�ΡC�H�ͳ����`�l���ʰϰ�d��q80��120
m2�C�H���R������v(resting ventilation rate)�O�j���C����6
���ɪŮ�A �ޤJ�j�q�~�ӷL�ɩM�Ů𤤪����j�]�t�i��b���f�r��ͳ��C�L�ɪ��H�����M�̪��j�p:
�D�`�p�����ɪ��l�J���ɭP�q�L�p�۩Τ��ިt�Ϊ��l���C���|�j��1 ��4 �gm���p�p�w�|�I���b�p��D�C���j���L�ɤ���i�J�I�l�t�ΩγQ�H���b�W�I�l�D(��1A)
�C
���h���J�D���m����]�A�����ê���d�I�l�D����ǬV�C�I�l�D�Q���H���֤h�]�A�֤�ӭM(ciliated
cells)�A �H�G���c�ӭM(mucus-secreting cells)�M��(glands)���л\(��1
B) �C�~�ӷL�ɦb��}�ΤW�I�l�D�Q�H�G(mucus)�ҧx�A �B�^����Y�Χ]�U�C�~�ӷL�ɱq���C���I�l�D�Q�W�ֲӭM(epithelial
cells)���֤��ʴ��W�C�b�ʥF�֤���H�G���ͪw�U�A �ѥ����ӭM(macrophages)�t�d���a�L��(��1)
�C
��J�D�ӭM�����X(Binding)
�y��ʷP�_�f�r���D�n�ؼЬO�I�l�D���W���W�ֲӭM�C�p�G�f�r�P�����O�s�b�M�u�@���A
�o�ǤW�ֲӭM�]�\�O�����ǬV���C�]�ӡA �f�r�P�����O���M�w�ʪ��ʦV(tropism)�C���O�A
�o��²�ƪ��ҫ��g�`�O�����H�����f�r�}�ʡA �]���P�����b�J�D�����G�@���w�[��f�r�V�ʤ��G���s�C
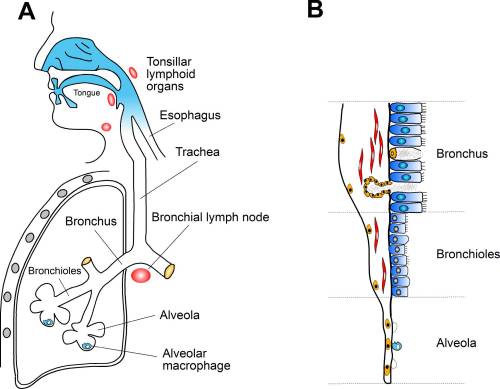
��1 . �y��ʷP�_�b�I�l�D�i�J���a�I�C
(A) ��ܤH����D���ѭ�M�\��c�y�C�y��ʷP�_�����ǬV�W��D�M�b����(bronchus)�M�Ӥ��ު�(bronchioli)�֤�ӭM�C�ް_���{�ɺ�X�g���]�A��ު�(tracheitis)�B���ު�(bronchitis)�B�Ӥ��ު�(bronchiolitis)�A
�M���ުͪ� (bronchopneumonia)�C��A�����K�̤����ѲO�ڵ��u��D�}�l�C(B)
�I�l�W�֥Ѥ@�h�H�G(����)�A �֤�ӭM(���ީM�Ӥ���)�A �M�ͪw�������ӭM(�ͪw)�W���S�O�˳ƫO�ñ���ӨӪ��f��ͪ��C
�b�y��ʷP�_���ǬV�W�A �f�r��y������(hemagglutinin)(HA)
�P�����s����m���ݳs����J�D�ӭM�������s���bֺ�T(sialic acid)�h�W���ĥb�ſ}
(galactose)(Weis 1988) �CHA�s����m���Y�ǰϰ�b�y��ʷP�_�f�r�ȫ������O���׳Q�O�u��(Daniels
1984)�C�J�D�i��i�H�ѴX�Ӿ����s��:
(1) �S���ʧK�̤���(specific immune response)�M�S����IgA
���骺���c���A (2) �D�S���ʪ�����(unspecific mechanisms)�A
Ĵ�p�H���֤�(mucociliary)���M�����H�J��(mucoproteins)���Ͳ���s����f�r��y�������A
��(3) �J�D�P����(ֺ�T�h��)����]�h�ˤơA ���׳Q�O�s�b�P�˪������A �����P�b���M�H���P��������(Matrosovich
2000)�C ���G�A ���f�r�ݭn�b��y�������P�����s������m�i����ܡA �h�q�L���M�H������V�J�D����ê�C�b�ް����W�A
ֺ�T�h�ĺ����h�Ω�(polymorphisms) �M��H�ó����b�ſ}�b��´�W���s�X�Q�@���F(co-expressed)�C�ҥH�A
�@�ǬV�ó��M�H���y��ʷP�_�i��o�ͦb�ީM��e�\���äH�ئb�@�Q�ǬV��(co-infected)�ӭM�ܭ�S�ʪ���]���աC�̪�A
����ܬY�dz��y��ʷP�_�f�r�b�H�M���W��s���줣�P���ؼвӭM(Matrosovich 2004)
�C�o������q90 �~�N�������H���y��ʷP�_�q�V�b�����a�ǬV��H���X�Ӯץ��[��CH5N1
�M�@�Ǩ�L�ȫ�(subtypes)�y��ʷP�_A �f�r��s����b�H���W���P���� (Olofson
2005) �C
�M�y��ʷP�_�f�r���s���@�˭��n���O�q���b�H�D�ӭM�W�s����m������(Cleavage)�C���άO�f�r���g�i�ĞJ�ï�(neuraminidase)���\�ਤ��(Chen
1998)�C�y��ʷP�_�f�r���r��(virulence)���M�g�i�ĞJ�ï��P��y���������ݮe�ʡC�@�Ӥw�g�b��y�������i�����(mutations)���@�r�f�r�ݭn�b���g�i�ĞJ�ï��i����ܥH�O����r��(Baigent
& McCauley 2003�A Hulse 2004)�C ���G�A �o�{�ﯫ�g�i�ĞJ�ï��ܤƾ�(inhibitors)�������ĩʪ��y��ʷP�_�f�r���A���O�M�r�ʬO��֤F��(Yen
2005)�C
���ӭM���M�f�r�ѯf�r�P����w�Q����a�æC(juxtaposed)�A
�ƦX��O�Q�ӭM�����](endocytosed.)�CH+ ���l�i�J��M��p�M(late endocytic
vesicles)�@���@���Ͳz�Ǩƥ�A �M��Ĥ�(acidification)�����C�b�ĤƤ��A
�f�rHA �i��@�ӧκA�����s��z�H�Ͳ��ĦX���D�J��(fusiogenic protein)�CHA
�������ϰ�(loop)���������骺���̲ױa�ӯf�r�M�M��p�M������[����H�K�ĦX�i�H�o�͡C���\�f�r�ֿ}�ֻĪ������ӭM�褺�A
�Ϧb�ĩʭM��p�M����H+ ���l ��M2 ���l��D���J�f�r�����C���G�A �b�f�r�M�M��p�M��
���ĦX����A �]�˸ѦbM1 �M�ֿ}�ֳJ�ս�ƦX��(ribonuclein complex)������C���P�ױӷP
���ۤ��@�ΡA �f�r�ֿ}�ֻıqM1���}�C�f�r�ֿ}�ֻĵM��H���T�C(ATP)�̿�覡�Q��J�ӭM�ֶi�����(transcription)�M��Ķ�@��(translation)(Flint
2004)�C
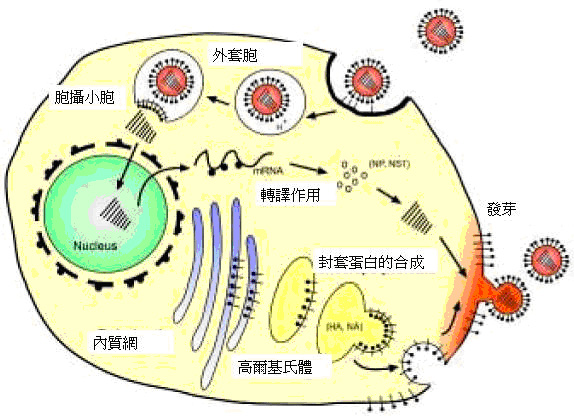
��2: �y��ʷP�_A �f�r���ƻs�g���C �f�r���s���M�I�J�A�H�M��p�M���ĦX�M�f�r�ֿ}�ֻĪ�����A
�b�ӭM�֤����ƻs�A ���c�M�ʮM�J��(envelope proteins)���X���A �f�r�o�ީM�����ǬV�F�~���W�ֲӭM
(�qCox & Kawaoka 1997��}��)�C
���n���ƻs�b���̵o��?
�ӭM�J�ջï�(proteases)���ݸg�`���ίf�r�J�ս�Φ������ӴI�ǬV�ʪ��f�r�L�ɡC�]�ӡA
�t�~�i�i�J�P�������]���i�M�w�f�r�ƻs����m�C�b�H���譱�A �y��ʷP�_�f�r�ƻs�@�������I�l�D�W�M�U�����W�ֲӭM�C�o�O�ѩ��(serine)�J�ջï��A
�سJ�էJ��(tryptase Clara)�A ���������ܡA �ѫD�֤���ޤW�֪��J�ԲӭM�Ҥ��c�C�Q�b�ƪ��ï����βӭM�~�L�ɤ����h�`����HA
�쪺�e��HA0�M���Ưf�r����HA �O�L�̨��P�V�C�@�ǰ��@�r�ʳ����y��ʷP�_���~�ءA �M�ӡA
�]�t�bHA �i�Q���Ϊ���m����]���J�ɭP�g���M�s�b�J�ջï�(ubiquitous protease)�B�z�C�o�]�\�ɭP�w�ק�L���V�ʤΡA�t�~�ƻs����m�b�ʪ��M�H����(Gamblin
2004 ) �C�b�H���W�������y��ʷP�_��´�}��(H5N1)�S���@�ӦX�A���w�q�C�b�@�ӮץA
�f�r�ֿ}�ֻĥѤ@�Ӥ�����E�X�Q��������A �Q�˥X�s�b��͡B�z�M��Ŧ�A ��������(positive-stranded)
�f�r�ֿ}�ֻġA �����f�r�ƻs�A �w���������ͩM�z��(Uiprasertkul 2005)
�C�]�ӡA �b�H����H5N1�f�r�ƻs�]�\�Q����I�l�D�M�p�z�D���P��b�䥦���Űʪ��M�������w�Q�ҩ���������(disseminated)�ǬV�C
�ǬV�f��˦b�J�D�Ǽ�?
�@���y��ʷP�_�w���IJv�a�ǬV�I�l�W�ֲӭM�A �b�X�p�ɤ����A
�ƻs�K�|���ͳ\�h�f�r�C�ǬV�ʪ��ɤl�q�W�ֲӭM���ӭM�������u���a�Q�����ͮ�D�A �o�ӹL�{�s�o��(budding)�C�]�F�~�ӭM�����t�ǬV�A
�o���Q��f�r�b�ͤ����ֳt�Ǽ��C �Ѧ۵M�o�ͪ�������bHA���Φ�m�����ܡA �o����ۦa�v�T�y��ʷP�_���V�ʩM�P�f�ʡC���G�A
���i�H�Ѩ䥦�ӭM���J�ջï����ѧO�C
�o�����쬰����b����\�h�Q�P�V�����y��ʷP�_(H5N1)���H�A
�����D(gastrointestinal)�B�x�M��Ŧ�A �M�I�l���g���M������o�DZw�̪��f�r�|��ѹ����ͯ��g�@�r(neurovirulent)
(Park 2002). �o�ǯg���O�_�_�]���Ǽ�(hematogenic spread)�ΤϬM�f�D��(non-pulmonal)�Φ����f�r�i�J��J�D���M�O�����Ԫ��C���O�A�bNA
�̪��ܤƤ]�i��A ���@�����A �����y��ʷP�_���o�ͦb�\�h��´�W��(pantropic)����C�Ҧp�A
���۹���Ǭy��ʷP�_��WSN/33�~�ءA �Ĥ@�ӳQ�������H���y��ʷP�_�f�r������A ���P��h�ƤH���y��ʷP�_�~�ءA
��b�պޤ��b�S���[�H�سJ�զӽƻs�C�b�o�f�r�A �@�ӽX�����R��(in-frame deletion)�h���bNA�ݰ�46
(residue 46) �W���ޤ�(glycosylation)��m���\���g�i�ĞJ�ï��s���M�j���M�߯���(plasminogen)�C�o�ɭP�a��ʴ��M�s�b���J�ջï��e���ܦ����@�סA�OHA�����μW�[�C�o�Ǭ�s���G���X�@�Ӭ�����y��ʷP�_A
�f�r�γ\�䥦�f�r�b�H�����ܱo���P�f�ʪ��覡�C(Goto & Kawaoka 1998)
�C���쪺�O�A ��]���ت�1918�~����y�檺��Z���y��ʷP�_�f�r(H1N1) ����s ���S�FNA
�ǹF��HA�i���ʪ��t�~����i��P���f�r�ƻs�M��r�ʬ��� (Tumpey 2005) �C
�ש�A �ʪ���s��ܡA ���ت�����(site of inoculation)�i�M�w�y��ʷP�_�f�r���b�J�D���Ǽ����u�C�Ҧp�A
�ݤ��ʥզ�y�� (neutrotropic) NWS�~�ظg�������q�L��� (hematogenous)�Ǽ������츣���A
�����f�r�Q���ئb��l���A ���̯�q�L��ı���g��(sensory neurons) ��F�������g�t��(central
nervous system) (Flint 2004). ��̡A �b����H5N1 �f�r���w�Q����(Park
2002)�C
�J�D�̪쪺�����O����?
���M�@�خɱ`�o�ͪ��e�f�A �M�@�ʪ��g�ʼҦ�(specific
inflammatory patterns) �ΧK�̤����ո`�Φb�H���y��ʷP�_���A�ӭM�f�ܪ��o�f��z�O�å������Q�A�ѡC
���y��ʷP�_�O�@�ش����ʯe�f�����i�A�h�O�Ӧ۰ʪ���s���ҾڡC�M�ӡA�o�˪��f�z�Ͳz�Ǽҫ��]�\�b�H���W���M���P�C
�ӭM�E��(cytokines) �M���f
�@�ӥD�n���D�O�W�b�I�l�D�@�ئa��ƪ��ǬV�O��˥i�ɭP�o���Y�����W���g��(constitutional
symptoms)�C �b�\�h�䥦�ǬV�f���A �D�M�@�ʤΦ��A���O���K�̤�����y��ʷP�_���{�ɯg��(signs)�M�g���ζǬV�᪺�����۷��j���^�m�C�o�ǧK�̾���i�ɭP�a��ʨèt�Ωʪ��@�ΡC�S�O�O�b�K�̨t�Τ����A
�b�ǬV�H��A �ѩI�l�H���W�֩M�K�̲ӭM���t�a�Q���ͪ��ӭM�E���A �O�E���ӭM�a��ʪ��E���A�C�ͤƿE��(chemokines)�O�@�����K�̨t�βӭM���X�ƪ���(chemoattractants)�O�ӭM�E�����@�Ӥl��(subset).
�Ҧp�A �y��ʷP�_�ǬV�b�H�����߲ӭM��(plasmacytoid)�M����𪬲ӭM(myeloid
dendritic cells) ���ɤ@���ͤƿE�����c���{�Ǯe�\���P���K�̨��ʾ�(effector)�Q��զa�l��(Piqueras
2005�A Schmitz 2005)�C �̭��n���ӭM�E�������]�ʭP����(endogenous
pyrogens)�M�Q���J�b���f���o�f��z����: IL-1Α/Β�A
TNF Α/Β�A IL-6�A�z�Z��(interferon)(IFN)
Α/γ�A IL-8�A �M�����ӭM���ʳJ��(macrophage
inflammatory protein)(MIP)-1Α�C
�o�DzӭM�E�����@�j�����b�g����ʩΦ۵M�ʥH�P�V�y��ʷP�_���H������|�~�����Q�d�X
(Brydon 2005) �C����ij���o�ǧ����ʩΨt�ΩʳQ�Ͳ����ӭM�E���A ���ۥ~�]�ʭP����(exogenous
pyrogens) (�p�y��ʷP�_) �P�]���ӭM���ۤ��@�ΡA ��F�������g�t�ΡC�b�U�C���A
���@�p�d��A �٦�ޯ��O�ݾ��x(Organum vasculosum laminae
terminalis)�A �䤺���@�ӳQ��֪��帣 �پ�(Blood-Brain-Barrier)�M���\�P�����q�L�C�b�o�Ӧ�m�A
�H�̿ྯ�q(dose-dependent)�覡�A �L�̻��ɫe�C����(prostaglandins)
�A �S�O�O�e�C����E2 ���Ͳ��C�o�Ƕǻ���(mediators)�W�[���ž��T�I(thermostatic
set point)�MIJ�o������Žո`����(thermoregulatory mechanisms)�W�q��šC��ڤW�A�b�H�W�Q���Ϊ��ӭM�E���P�y��ʷP�_�ǬV���Y���ʨèS���Q���p�A
���ɦV�̪���]�h�ĩ�(pleiotropy)�M�b�H����|���������ۤz�Z�C�ӭM�E���������ʤ]�\�b�y��ʷP�_�~�ةέ��餧���������P���a��C�y��ʷP�_�ǬV�H����1997�~��H5N1
�~�سQ��ij���Ħa���ɦ�NS��]�������e�o��(pro-inflammatory)�ӭM�E��(�S�O�OTNFΑ)
(Cheung 2002�A Lipatov 2005�A Chan 2005). ��s�ت��O�b���{��L���ɲӭM�E�����f�r�ե����:���ױq�Q�ǬV���ѹ����͡A
�άO�q�y��ʷP�_�f�r�X�������o������(double-stranded) (ds) �ֿ}�ֻķ��`�g�J�ѹ�CNS
�߫ǮɡA �O�o���ʪ��C�����̦���A �o�˪�dsRNA �q�Q�ǬV���ӭM�Q�����~�Ө�E�ӭM�E���Ͳ��C�̪��s��ܡA
����P����������(dsRNA-sensing Toll-like receptor) (TLR)
3�Q���F�b�ͤW�ֲӭM ��TLR3 �����ѻP��I�l�W�ֲӭM���K�̤��� (Guillot 2005�A
Akira & Takeda 2004)�C���쪺�O�A�@�ӤH���P�ͭѨӹ�ܬy��ʷP�_�K�̤������}�l�A
�ۤ��TLR 3�d�XdsDNA�A ���M�ʤ��ȩ�q�LTLR 8����O����ֿ}�ֻ�(single
stranded RNA)�C�f�r�L�ɥi�ೣ�O�P���ʪ��A �]�Q���h�ֿ}�ֻĦ��]�A�f�r�o�סA
��y�������A �M���g�i��?�ï����L�r�� (virosomes) �]�\�ɭP�o�N�C�M�ӡA�ӧO�f�r�ե�O�D
�P���ʪ��A �j�������������f�r�̭]�ಣ���y��ʷP�_���g�������椸(subunit)�̭]�h����(Brydon
2005) �C
�I�l�g��
���ިt�ιL�פ���(Hyperreactivity)(Utell
1980�A Little 1978)�A��ê�D�n�p��D(Hall 1976)�δ�z���X���e�q(Horner
1973)�O�b�y��ʷP�_�ǬV���q���C�L�פ����M���ު�ê�]�\�����@�q���ɶ��A �S�O�O�b�L�ӯe�f(Kondo
& Abe 1991)�A �M�]�\�O�e���ʲӭM�E�����p(pro-inflammatory
cytokine profile)�H�z�A�ɭP�@���B��ʭP�ӭ�(aerosolized allergens)����O
(Tsitoura 2000) �C
�b�H���y��ʷP�_�ǬV�A �Y���ͪw���g���D�n�f�r�ͪ��A �O�u�����C���q�`���X�U�M�W���I�l�D���������o���P�֤�ӭM�l���B��hyperemic
�ΥX��ʰϰ�b�z����(hyaline membranes) �B �ݤ��ʩM��ֲӭM�����J(Yeldandi
& Colby 1994) �C �P�D�n�f�r�ͪ����A �ӵ߭��ƶǬV(bacterial
superinfection) �O�b�H���y��ʷP�_���`�����M����ޭP�Y���o�f�v�M���`�v
�S�O�O�b�~�������H�C�Q��{���X�Ӧ]���A ������I�l�D�ӵ߶ǬV���I���W�[�A �o�]�A�W���W�ֲӭM�l�˻P�W�ֲӭM��ê���}�a(Mori
1995)�A ����H���֤M��(Levandovsi 1985)�A ��i�ӵߪ��H��(McCullers
2002)�A �M���ʥզ�y(neutrophils)���\�����(Abramson 1986�A
Cassidy 1988)�C
�ӭM�f�A
�H���y��ʷP�_�ɭP�����ӭM�f�A�A �D�n�b�I�l�D�W���W�ֲӭM�A
�ɭP��ʪͩM��D�e�f�C�y��ʷP�_�f�r�b�I�l�D���ǬV�M���f�r�ƻs�ɭP�ӭM�l�˳Q���ɱH�D�ӭM�J�ս��X���U���ո`(downregulation)
(Katze 1986�A Sanz-Esquerro 1995) �òӭM�p�e�ʦ��`(apoptosis)
(Wiley 2001a)�C��̡A �S�W�ӭM�{���ʦ��`�A �O�@�t�C���Q�w�q���ӭM�ƥ�̲ɭP���IJv�a�M���ӭM�M���������C�ӭM�p�e�ʦ��`�i��Ѥ��P������IJ�o�M�H�κA�W�����ܹ�ı�A
�]�A�ӭM���[(cytoskeleton)���}�a�A �ӭM��M�֬V��(chromatin)�����E�B�u����@�Υ��ġBDNA�_�H�A
�M�̫�A �p�ӭM�����s�ɤl�W�� ��`�p��(apoptotic bodies) ���Φ��A
��Q�]���ӭMĴ�p�����ӭM�M�𪬲ӭM(dendritic cells)�M���C
�y��ʷP�_�f�r�o���ӭM�p�e�ʦ��`�O��Fas�ǹF����MFas�W�߫H���ǹF���A
Ĵ�pFADD/caspase-8 �ƦX��ѳJ�տE�Q R (Protein Kinase
R) (PKR)�ҧΦ� �A �}�l�@�����֮�ijJ���Q(caspase) ��y(cascade)�C
��IFN���ɤγQdsDNA�E�ƪ�PKR�O�b�\�h��`�~�|(apoptotic
pathways) �����@�����䱱��ե� (Brydon 2005)�C�@�����ӭM�p�e�ʦ��`���ĤT�����A
�y��ʷP�_�q�L�f�r���g�i�ĞJ�ï�(neuraminidase)�E���ܫ��ͪ��]�l���L(TGF)-Β�CNA�i��ѫP�iTGF-Β���Φܥ������D�Φ��E����b�b�ӭM������TGF-Β�CTGF-Β
�_�l�@�ӫH����y(cascade).�p�r���ɭP c-Jun N �ݳJ�ս�E�J(c-Jun
N-terminal kinase) (JNK) ���������ƫ��J�ս�E�J(stress
activated protein kinase)�A �y������]�������ƩM�e��`��]���F(pro-apoptotic
gene expression) ���W��j�X(upregulation)�C�o���~�|�A �P�p�J�ժ��u���齤�����A
�QĶ���bPB1 �J�ս誺�@�ӿ��+1�\Ū�ج[ (+1 reading frame) (Chen
2001)�A �w�Qĭ�t�b�O�ڲӭM�p�e�ʦ��`�ί�����b��ʶǬV�������O�ڲy��֯g(lymphopenia)�C
�Ͳ�´���˫�ӨӪ��y��ʷP�_�f�r���ǬV�w�P�ӭM������O(oxidative
stress)�A ���ʮ�(reactive oxygen species) (ROS)���Ͳ�
�M�v�Į�ƪ��X���ï�2 (nitric oxide synthetase-2) �����A
�ɭP���������r�ʴ�𤤶���(intermediates)���Φ��C�M�ӡA �ܮA �b�պޤ�������(bronchiolar)�ӭM�u�����ӭM�p�e�ʦ��`�u�O���ܤ֪��@�ΡC
H5N1���ǬV�g��
�����y��ʷP�_�O�����ǬV�f��A���~�جy��ʷP�_�f�r�ҳy���C�����A
�Ҧ����P�f���z�o���Φ��O�Ѭy��ʷP�_�ȫ�H5�MH7 A�f�r�ҳy���C���e�����y��ʷP�_�b�H��(H5N1)
�O�_�M�H�W���쪺�ӭM�f�A�@�ˬO�����ơC�u���ܤ֩v�Y���έP�R���ץ�Q��s�C���O�A �L�g���ηũM�g���ʪ�(symptomatic)�e�f�O�i�(Buxton
Bridges 2000�A Katz 1999) �è�o�Ͳv�]�\�Q�C���C
H5N1 �y��ʷP�_�b�H���̦@�P������g���O���N�B�M�b���Ǧ���|���w�̦��ͪ��B�|��(pharyngitis)�B�p�z�g���B������(conjunctivitis)�A
�M��ʸ���(encephalitis)(Yuen 1998�A Tran 2004�A Yuen
& Wong 2005).�C������x���O�ͪ������~�f�H�g�`�i�@�B�F�o�i����ARDS���e�f�C�bH5N1
�y��ʷP�_���P�R�f�ҡA ������������g�Ըs(Hemophagocytic Syndrome)�Q�y�z���@�Ӭ�X���S�I�C
�b�ͯf�H��´�X���ͭM�l�˩M�ӭM�����ֺ��ܩʤ��~�A �ͥ~�� ��(extrapulmonary)�����J�w�Q�y�z���s�x�ʨx�����p�����a��(extensive
hepatic central lobular necrosis)�A ��ʵǤp���a�� (renal
tubular necrosis)�òO�گʥF(lymphoid depletion) (To
2001)�A ���M�b������S���f�r�Q�o�{�A������E�X�����(Reverse Transcription-Polymerase
Chain Reaction�M�K�̬V��]immunostaining�^���յo�{�i���Ѫ����ս�l-2
�P����(interleukin-2 receptor)�B�ս�l-6 ( interleukin-6)�M�z�Z������(interferon-gamma)���W�[�F�C�t�~�A
�~�F�a���]�������k(tumor necrosis factor-alpha) mRNA
�b�䥦�H�H��H5N1 �y��ʷP�_���Ͳ�´���f�Ҥ��Q�o�{(Uiprasertkul 2005)
�C
�P�H��H1N1 �f�r���(Hayden 1998)�A �q1997
�~������H5N1�~�ؤw�Q��ij�䦳�ĤO�a��NS��]�������ɫe���ʲӭM�E���]�AIL-10�BIFNΒ�BRANTES�BIL-6
�M�S�O�OTNF (Cheung 2002�A Lipatov 2005�A Chan 2005)�C�o�Ǭ�s���@�̰��]�A
�b�P�R�ʤH���ǬV�ʳ����ȫ�H5N1���A �̪쪺�b�I�l�D���f�r�ƻsIJ�o�Ѥ@�ө�����������g�Ըs�ҽ����ƪ��ӭM������(hypercytokinemia)
�]�\�O���O��q�`�H���l���H�~���y��ʷP�_A H5N1�ǬV���o�f��z (To 2001)�C�bH5N1
�����y��ʷP�_���P�R�f�Ҥ��å��o�{�ӵ߭��ƶǬV(To 2001)�C�o�[��]�\�O�o�dz��Y���������P�R�f�ҵ��G�������A
���]�a�S���e�\���ƶǬV���o�i�C
�y��ʷP�_��˳Q�Ǽ����L�H��?
�I�l�ʶǼ����M��a�f�r����L�ɩM�B�誺�Ͳ��C�b���ܩM���`�I�l�����A
�B��|�Ͳ��C�q��Ĭy�X�n�D���Q���M�p�G�ǬV�s�y�F�@�ػ���c���A ���K�i�Ħa�y�X�C�Q���Ͳ�20�A000
�p�w�P�u���y�¤���X�ʤp�w�Q�X�C�̤j���p�w�U�^��X�̤������a���C�Ѿl�p�w���ʪ��Z�����M�b���̪��j�p�C���|1-4�gm���p�w�]�\�����O�����d�Ψ�F�U�I�l�D�C����ʬy��ʷP�_�ǬV�����@����ܡA
�p�p�w�b���ު��l�J����j�p�w���J�W�I�l�D�β��y���������ج��h�C (Alford 1966�A
Little 1979�A Bridges 2003). �p�G�f�r�b�����ǬV����U�I�l�D�ƻs�A
�o�|�ɭP��p�B������h�f�r�ΧǬV�O���p�w�X�{�A �]���S���ʧK�̺ʱ��t�Τ��M���إߡCH5N1�q�ʪ���H���ǬV�]�\�H���P���覡����(�M����)��IJ�Q�ǬV���V�b�C
�������v�O�ɭP�y��ʷP�_A �y����z�o�ҥ��ݪ��C�ҥH�A �V�Ѭy��f�b�ڬw�M�_���w�]�\�]����[�h��Z������IJ�M�r�d�b���q�����ж��C�y��ʷP�_�f�r���A���O�O�j��:������G���ͦs����O�b���C���۹���שM��C�����ҷū���(Hemmes
1960) �C�����y��ʷP�_(H5N1) �]�\���ӾA���p�w�ǬV: �����O������(Chotpitayasunondh
2005)�A �z�פW�b�y��ʴ������y�����֪��\�h�H���@�ֶ}�l�V�f�C�p�z�ƻs�M�g���b�I�l��ܦܤ@�ӬP��(Apisarnthanarak
2004 �H�e)�A ���\�S���ʧK�̤����b�ǬV�ʤp�w�Ǽ����e�_�l�t�ܡC���G�A�b�����y��ʷP�_����|�ƻs���֦b�H���y��ʷP�_�o��(Peiris
2004) ���f�r�ƻs�O�Q����(Beigel 2005) �C����{�b�A H5N1�b�H�����ǬV�O�u����(Buxton
Bridges 2000�A Ungchusak 2005)�M�۷��L�Ī��C�`�Ө����A �����y��ʷP�_�f�r(H5N1)
�ڱ����H�ǤH���ǬV�M�̫�F��O�_�������Ĥް_�y��ʩΤj�y�檺�P�V�v�n�X�ӶǥN(passages)�C
�K�̾�
�y��ʷP�_�ɭP�J�D��ʷP�ǬV�M�_�l�K�̤�����y(cascade)
�E���X�G�K�̨��m�t�Ϊ��Ҧ������C�j�h�Ƴ̪쪺���ѩʤ����A �]�A�ӭM�E������(IFNΑ/Β)�A
�J���ݤ��ʲɲӭM(neutrophil Granulocytes) �Φ۵M����ӭM(natural
killer cells) (Mandelboim 2001�A Achdount 2003)�A
�M�ӭM���Ƨ@�ΡA �t�d�_�l����{�ɯg��(�Ѩ��W��) �C���ѩʧK�̬O��ѩʧK�̤���(adaptive
immune response)���n���A �����A ����̪쪺�f�r�ƻs�M�ܭ�˸��A �βĤG�A
�]����ѩʧK�̤������ܭ�M�@�ʲO�ڲӭM�O�Q�b���̻P�f�r�����@�δ����A �Q���ѩʧK�̨t�βӭM���ɪ���P��E���l(co-stimulatory
molecules)�ҿE�� (��3)�C�y��ʷP�_�f�r�A �M�ӡA Ķ�X�b�D���c�ʳJ�ս�1(NS1)
������שM���IFNΑ/Β �����CNS1�i��i�H�j���f�rdsRNA����ѲӭM�P�����ѧO�o�M�I���l�_�h�|IJ�oIFN
Α/Β ����(Garcia-Sastre 1998�A Garcia-Sastre
2005).��o�ʧK�̤���(adaptive immune response)�ݭn�Ƥ�~���Ħ��������U�]�t�f�r�Ǽ��A
�n�����f�r�A �M�̫�إ߰O�Ф����H�T�O�i�����ʩ�ܦP���f�r���A�P�V�C�b�y��ʷP�_�ȫ�����t�O�@�ܤֳQ��ı��åB�ǬV�ڥ��L�k���P���ȫ���A��B���������O�@(Treanor
2005). �y��ʷP�_�P�V�ɭP�t�ΤΦa��ʧ��� (��G�K��) (humoral immunity)�A
�ìr����T�ӭM(cytotoxic T cell)���� (�ӭM�ʧK��) (cellular
immunity)�A �C�ӳ���q��ʶǬV���_���M�A�P�V����ܤ������n�C
��3 . ��y��ʷP�_�f�r�ǬV����G�M�ӭM�ʧK�̤����C��G�ʧK�̨t�Ϊ�����]�AB
�O�ڲӭM(��)�A �b�P�y��ʷP�_���@�ΥH��A ���Ʀ����c���骺��߲ӭM�C�ӭM�ʤ���(�k)
�H�ܭ�e�{�q�LMHC I (�¦�) �åBII (�Ŧ�) ���l�Ѿ𪬲ӭM�}�l�A �M��ޭP���Ƨ@�ΡB�ܭ����T
�ӭM���E�W�M����(CD4 ��CD8) �C�o�DzӭM����ʧ@���ӭM�@�Ϊ����a���U�A ����ӭM�E���A
�ζǹF�ӭM�r��(cytotoxicity)���ѧO�ܭ� (Adapted from Flint
2004)�C�S���b���Q��ܪ��O�Ѭy��ʷP�_���ɪ��ӭM�O�ЧK�̤����M���P�Φ������ѩʧK�̤O�C
��G�ʧK��
����(�pIgG�A IgA)�O��B �ӭM�o�i�̫ᶥ�q����߲ӭM�Ͳ��A
�ݭnB �ӭM�w�ѧO�ܭ�Τw�QCD4 T �ӭM�MT �ӭM�l�ͪ��ӭM�E��(��3)��E�C�PT
�ӭM���P�A B�ӭM�i�H��즳�Φ��ѧO�ܭ�C�ܭ�S���ʱq��]���H�N����s�X�b�ӭM�����K�̲y�J�հ��i�ܲ�(hypervariable)�ϰ�X�{�A
���ӭM���M�b����C�M��A ������(Native) B �ӭM�i�J�`���A �q�L��G�M�O�ڸ�(lymphatics)�ܲ�´�M�O�ڸ����x�C�b�O�ڵ�(lymph
nodes)�A ������(native) B �ӭM�ѥ��̪����������ѧO�P���ܭ�A �ܦ����E���A�qIgM
�ഫ(switch) ��IgG �Ͳ�(�����ഫ) (class-switch)�A �W�[���̪��K�̲y�J�կS���ʩM�˦X�O(affinity)�A
�η��ӭM�~��b�ӭM�E�����v�T�U�����A ���Ʀ���߲ӭM�ΰO��B �ӭM�C��IgA �Q�B����W��D���H���W�ֲӭM�A
��ΥH���M�βM���f�r�ǬV�A IgG �D�n�t�d�O�@�U�I�l�D�C (Palladino 1995�A
Renegar 2004)�C
�y��ʷP�_�ǬV�ɭP��y��ʷP�_�}�J��HA�A NA�A M �MNP
�J�ս誺����t�ΩʥͲ��C�Ҧp�A HA �M�@�ʧK�̲y�J�աA �]�AIgM �BIgA �MIgG�A
�X�{�b�f�r���ؤ���P�����C��NA�o�i�P��y�������ܤƩʧ���(hemagglutinin-inhibiting
antibodies)���o�i����C�b�ǬV�H�᪺4-7 �ӬP�������A �b����w�w�q�K�F��p���A
�H�ۥ�í���U���C�b�ǬV�H��A ����i�O�d�X�~�A ���ިS�����_���S(re-exposure)�C��HA����P�O�@�K���P���f�r���ǬV�Τo�e�f�A
�åB���ɤ��M���骺�O�̭]�K�б��ط������@�ӥD�n�ؼСC��MHA �ܤƺw�w�q1:40 �Χ�j�A
�Φ�M���M�w�w�q1:8 �Χ�j�A ���i�O�@�K���ǬV�C�b���Ѫ��H�餤�h�ݭn����������A
�~�i�����O�@ (Treanor 2005) �C
�P��HA������A ��NA���餣�त�M�f�r�P�V�A ���O�ϦӴ�֯f�r�q�Q�ǬV���ӭM���IJv������
(Johansson 1989) �C�o�O�]�����g�i�ĞJ�ï��A�}�����̪�Φ����L�ɪ��ӭM�P������ֺ�T�h�Ī��ݰ�C��NA
����i�H�O�@�K���e�f�ε��G��֯f�r��������O�M�Y�����g���C���H��ij�]���ۦ����@�ι�y��ʷP�_A
��M2 �J�ս誺����A ���M�`�A�ӻ��A ����鷺���ܭ�O�D���M�ʪ��A ���|��֪������M�ݨӤ���R�����@�K�̪��@�Ө���C
�H���K�̤�����y��ʷP�_�A �̷Ӧb�i���q�o�����c���A �H��HA
��IgA �MIgG1���S�x�C�H����HA IgG �����P���M�������K�����p�A �����q�t�ιj�����Q���X���A
��IgA �O�����a�Q�Ͳ��C��s��ij�A��A�P�V����ܬO�D�n�a�ѧ����ʳQ�Ͳ���HA �M�@��IgA�ҶǹF�A���M�]�\�O�MIgG������(Renegar
2004)�C�p�G���R�����@�סA �H���γ�W�t�Ωʧ���i�H�O���@�ʪ��A ���̲z�Q�O�@�o�ͩ��M�M�����æs��
(Treanor 2005)�C����b�K�̤���y��ʷP�_����ʬO�Ѥ��M�f�r�A �Υ˸ѳQ�ǬV���ӭM�f�r�q�L����Χ���̿૬�ӭM�r�ʧ@��
(antibody-dependent cellular toxicity)�C
�b��ʯf�r�ǬV�f�r�������f�r���D�H�O��ӦۦP�˯f�r���ǬV�����M���K�̤O�C�M�ӡA
���ަ����D���K�̲M���A �Ѭy��ʷP�_�f�r�y������ʶǬV�@�A�o�͡C�o�O�]���y��ʷP�_��ܤ@�i��ʪ����c�Ψ��e�Ԧb�������c�J�ս�̦��\�h���Ĵ��N�ӨS�����h�����ǬV�O�C�Ҧp�A�s�����lHA��ֺ�T�h�ķP�����A
�t�d�f�r��ؼвӭM���i�J�A �åB�O�@�Ӭ����M����M�ӭM�r����T �O�ڲӭM�D�n�ؼСA �i�ܳs��K�����O�C�o�K�̿�ܩΦh�ˤơA�q�ƻs�����~�X�{�A
�ɭP���LHA�H�ɶ����ܲ��Ӥ��\�f�r�k�פH���K�̤��� (�ܭ�}��)�]Antigenic drift�^�C�o���ܰʬO�y��ʷP�_�C�~���y��ʶǼ�����]�M�b�C�~�y��ʤ��e�ݭn�����Ʒs�̭]�C�ۤϡA
�b��]�թΰ�]�լq���A���X�έ��եH��A �]����]�����aĶ�X���s�����J�ս�A �ܭ�}���O�b�f�r�����J�ս�W���@�ӥD�n�ܤơC�C���V����ƻs�ɡA
�ܭ��ഫ�O�i��C�ۤϡA �ܭ�}���u��b�Y�DZ��p���U�o�͡A �O�۹�a�u���M�i��O�j�y�檺�@�ӭ�]�C
�ӭM�K�̤���
�𪬲ӭM�Q��ܦb�}�l�M�r�pT �O�ڲӭM��������t�@�ӥD�n�D���C���̬O�}���a���G������l�ͪ��զ�y�E���p�ձM�����ܭ��T
�ӭM������A �B��A �B�z�M�i�� (��3) �C���d�ҬO�A �s�b��ͤ����𪬲ӭM�q�I�J���f��ͪ����o�ܭ�A
�ܬ��E���A ���H�᩹�����ʤެy�O�ڵ�(Legge & Braciale 2003) �C�ܭ�˥��w�B�z�M�Q�T�w�b�𪬲ӭM�����@���ѥD�n��´�ۮe�ʽ�����(major
histocompatibility complex )(MHC)���l���Y���(Silver
1992). �b�O�ڵ��A �{�b�����𪬲ӭM���IJv�a�ѥ���@��T �ӭM�P�b�𪬲ӭM�������S���ʥ~��-�Y���-MHC
�ƦX��P����IJ�o�@�ӧK�̤���(Shortman & Liu 2002)�C�q�𪬲ӭM���f�r�ǬV�����]�ʧܭ�Q�B�z�γQ�i�ܳq�LMHC
I ���l��CD8 T �ӭM�C�~�]�ʧܭ�q�LMHC II ���l��CD4 T �O�ڲӭM�Q�i�ܡC�G�̾ܤ@�a�A
�𪬲ӭM�]�\�i�i�ܥ��̱q�Q�ǬV���ӭM������ܭ�A ���ಾ�ܭ�ܦb�O�ڵ����F�𪬲ӭM�A
�M��Ѥ@�ӵ{�ǥs��e�ܭ촣�e(cross-presentation)�_�lCD8 T�ӭM����
(Belz 2004�A Heath 2004�A Wilson 2006).�̪�Q�E����T
�ӭM������ʾ��ӭM�@�ΩM���~�ܦb�ͤ����ǬV�a�I�H�K�ǹF���̪��ܯf�r����(��3) �C
�b�P�V�_���H��A �K�̾ǰO�Ъ��A���ۦӨӦb���� �����H��P�˯f�쪺�ǬV
(Ahmed & Gray 1996)�C�O�ЬO�ѧܭ�S����T �ӭM�H����W�[���W�v�Һ���
�A �P������ (na?ve) T �ӭM����A ��֤F�@��E(co-stimulatory)�H�����ݭn�A
�M��ܭ�A��E(restimulation)�������t������ (Woodland & Scott
2005). �禳�ҾڶɦV��y��ʷP�_�S����CD8 �O��T �ӭM�b�H���̪��ֿn���a�I���S���ʡA
����ͦA�P�V�@���K�̩ʫO�@(de Bree 2005�A Wiley 2001b)�C�b�y��ʷP�_�ǬV�����A
CD4 �MCD8 �O��T�ӭM�ȸs�]subset�^�����A �M�ǹF����y��ʷP�_�f�r���A�P�V�A
�O�P�즸�P�V�f�r���M���D�n�̾aCD8 T �O�ڲӭM���@�Τ��P(Woodland 2003)�C
�t�@���n�S�x�A �Ҧp�b�y��ʷP�_�ǬV�A �OCD4 T �O�ڲӭM���UB
�O�ڲӭM���ͧ�HA�M��NA ����(��3) �C�bHA��CD4 T ���U�ӭM�ѧO���f�r������Ѫ��ϰ즳�O��ѳQ���ǧ����ѧO���ϰ�CT
���U(Th) �ӭM�]�\��P�i�f�r�S����CD8 �ӭM�r��T �O�ڲӭM�����͡CTh �ӭM�ܤ֥i�ھ��������̥Ͳ����ӭM�E���A
�i�@�B�Q�Ӥ���Th1�MTh2 �ӭM�A�C�b�ѹ��A �y��ʷP�_�ǬV�ɭP�@�ӱj�P��Th1 �����A
��Th2�ӭM�E���A (IL-4 �BIL-5 �BIL-6�A IL-10) ��Q�o�{�b�Q�ǬV�ʪ����ͳ��C�@���Ҿڪ����A
���@�K�̬O�ѹ�Th1�����Q�ǹF�C�b�y��ʷP�_�ǬV�A CD8 �ӭM�r����T �O�ڲӭM(CTL)
�qHA �Τ����J�ս�M �BNP�A �γQ�i�ܦbMHC �� I ���l��PB2�ѧO�f�r������Ѫ��ϰ�
(Treanor 2005) �C�ھڥ��̪��ܭ�S���ʡA CTLs �]�\�O�ȸs �S���ʪ��ΡA
�p�G�����ѧO�����ܭ�A ��e�s��e�a�P�y��ʷP�_A�����C�ʪ�����ϥαįǦ�CTLs�հ����S�F���̦b�ǬV�������W�ͩM�E���˦�(Lawrence
& Braciale 2004�A Lawrence 2005) �è�A�q�y��ʷP�_�ǬV�F�첬¡����O�C�M�ӡA
�L�̦b�y��ʷP�_��������O����a�ݭn�C
�b�H��T�O�ڲӭM�����F�̰��I�j���b��ǬV����14�� �åB�y��ʷP�_�S����CTLs
�������P���H�f�r�ƻs�������M��������֦��������p�C�O��CD8 T �ӭM�]�i�b�ﵽ�e�f�Y���ʩM�P�i�A�P�V�ɪ��_���R���@�Ө���C�̪�ʪ���s��ij�A
�b�̪��l�^����(recall response) �����X�ӵu�ȩM�b�ѭ�ǤW�Q�������������q�C�Ĥ@���q�Ѿn�b�ͮ�ު��O��T
�ӭM�ǹF(Woodland & Randall 2004)�C���n�a�A ���f�r�q�٬O�D�`�C�ɡA
�o�DzӭM���ǬV���Ĥ@�x�������� �C�ѩ��������ҭ���L�k��ǬV�ӼW�͡A ���̤]�\�ಣ�ͲӭM�E���H����f�r�ƻs�M�b�W�ֶǼ����C�ĤG���q�����A
�b�̪�ƤѪ������O�Ѩ��t�a�Q�x�J���ު��O��T�ӭM�ҶǹF�C�ĤT���q�O�o�ͦb���O�ڸ����x�O��T�ӭM�ܭ��X�Ϫ��X�i�C�o�ǰO�вӭM�H�X�Ѧb�O�ڸ����x�W�ͩM�u��b���ǬV���j��5
�ѫ�A �Q�x�J�����ءC(Woodland & Randall 2004)�C�o�DZq�ʪ�����ް_�������ҫ��O�_�A�Ω�H�������p�O�������C
�M�ӡA ���F��i���Ӭ̭]���p���A ���ݧ�n�a�A�ѧK�̤����������Ω�y��ʷP�_�ǬV�������Ī��O�СA
T�ӭM�������Ͳ��κ��@�C
����
�ڭ̬ݨ��F�y��ʷP�_�f�r�ǬV���ɭP���ʩI�l�f�g����ʵo�i�C�o�f��z�H�b�ͤ����f�r���t�ƻs�M���G���S�x�A
�y�������M�t�Ωʵo���βӭM�E������C�o�Ǩƥ�A �P��o�ʧK�̤����A ���U��֯f�r�t��A
�����f�r�A �MIJ�o�e�f�_���C��G�βӭM�ʧK�̤����A �ѶǬV�α��ػ��ɡA ���ѭ���Τj��������f�r�~�ت����[�ʧK�̨��@�C�M�ӡA
�y��ʷP�_�i�q�L�ܭ��ಾ�M�}���A �}�a�o�ǬV�ά̭]�l�ͪ��K�̡A �y���y��ʩM�j�y����z�o�C�N�ﵽ�A
�]�A��]�M�\���s�A �N���U�i�@�B�F�Ѿ��v�ʩM���e�y�q���y��ʷP�_�@�r�~�ت��o�f��z�C�Ʊ�o������H���ͳ��f�r�K�̨��m������ѩM���ײz�ѡA
�N�P�i��ܧ�n���v���M�蘆��y��ʷP�_�f�r���ܲ��A ����h���Ī��̭]�åi���t�a�o�i�Τ��G���@�ɡC
|
|
���
- Abramson JS, Wheeler JG,
Parce JW, et al. Suppression of endocytosis
in neutrophils by influenza A virus in vitro.
J Infect Dis 1986; 154: 456-63. Abstract:
http://amedeo.com/lit.php?id=3734493
- Achdout H, Arnon TI, Markel
G, et al. Enhanced recognition of human
NK receptors after influenza virus infection.
J Immunol 2003; 171: 915-23. Abstract: http://amedeo.com/lit.php?id=12847262
- Ahmed R, Gray D. Immunological
memory and protective immunity: understanding
their relation. Science 1996; 272: 54-60.
Abstract: http://amedeo.com/lit.php?id=8600537
- Akira S, Takeda K. Toll-like
receptor signalling. Nat Rev Immunol 2004;
4: 499-511. http://amedeo.com/lit.php?id=15229469
- Alford RH, Kasel JA, Gerone
PJ, Knight V. Human influenza resulting
from aerosol inhalation. Proc Soc Exp Biol
Med 1966; 122: 800-4. http://amedeo.com/lit.php?id=5918954
- Apisarnthanarak A, Kitphati
R, Thongphubeth K, et al. Atypical avian
influenza (H5N1). Emerg Infect Dis 2004;
10: 1321-4. Abstract: http://amedeo.com/lit.php?id=15324560
- Baigent SJ, McCauley JW.
Influenza type A in humans, mammals and
birds: determinants of virus virulence,
host-range and interspecies transmission.
Bioessays 2003; 25: 657-71. Abstract: http://amedeo.com/lit.php?id=12815721
- Beigel JH, Farrar J, Han
AM, et al. Avian influenza A (H5N1) infection
in humans. N Engl J Med 2005; 353: 1374-85.
http://amedeo.com/lit.php?id=16192482
- Belz GT, Smith CM, Kleinert
L, et al. Distinct migrating and nonmigrating
dendritic cell populations are involved
in MHC class I-restricted antigen presentation
after lung infection with virus. Proc Natl
Acad Sci U S A 2004; 101: 8670-5. Abstract:
http://amedeo.com/lit.php?id=15163797
- Bridges CB, Kuehnert MJ,
Hall CB. Transmission of influenza: implications
for control in health care settings. Clin
Infect Dis 2003; 37: 1094-101. Abstract:
http://amedeo.com/lit.php?id=14523774
- Brydon EW, Morris SJ, Sweet
C. Role of apoptosis and cytokines in influenza
virus morbidity. FEMS Microbiol Rev 2005;
29: 837-50. Abstract: http://amedeo.com/lit.php?id=16102605
- Buxton Bridges C, Katz JM,
Seto WH. Risk of influenza A (H5N1) infection
among health care workers exposed to patients
with influenza A (H5N1), Hong Kong. J Infect
Dis 2000; 181: 344-8. Abstract: http://amedeo.com/lit.php?id=10608786
- Cassidy LF, Lyles DS, Abramson
JS. Synthesis of viral proteins in polymorphonuclear
leukocytes infected with influenza A virus.
J Clin Microbiol 1988; 26: 1267-70. Abstract:
http://amedeo.com/lit.php?id=3045149
- Chan MC, Cheung CY, Chui
WH, et al. Proinflammatory cytokine responses
induced by influenza A (H5N1) viruses in
primary human alveolar and bronchial epithelial
cells. Respir Res 2005; 6: 135. Abstract:
http://amedeo.com/lit.php?id=16283933
- Chen W, Calvo PA, Malide
D, et al. A novel influenza A virus mitochondrial
protein that induces cell death. Nat Med
2001; 7: 1306-12. Abstract: http://amedeo.com/lit.php?id=11726970
- Cheung CY, Poon LL, Lau
AS, et al. Induction of proinflammatory
cytokines in human macrophages by influenza
A (H5N1) viruses: a mechanism for the unusual
severity of human disease? Lancet 2002;
360: 1831-7. Abstract: http://amedeo.com/lit.php?id=12480361
- Chotpitayasunondh T, Ungchusak
K, Hanshaoworakul W, et al. Human disease
from influenza A (H5N1), Thailand, 2004.
Emerg Infect Dis 2005; 11: 201-9. Abstract:
http://amedeo.com/lit.php?id=15752436
- Cox NJ, Kawaoka Y. Orthomyxoviruses:
Influenza. In: Topley & Wilson's Microbiology
and Microbial Infections, 9th ed., Collier
L, Balows A., Sussman M., eds., Edward Arnold,
London Vol.1, 1997: 385-433.
- Daniels RS, Douglas AR, Skehel
JJ, et al. Antigenic analyses of influenza
virus haemagglutinins with different receptor-binding
specificities. Virology 1984; 138: 174-7.
Abstract: http://amedeo.com/lit.php?id=6208680
- de Bree GJ, van Leeuwen EM,
Out TA, Jansen HM, Jonkers RE, van Lier
RA. Selective accumulation of differentiated
CD8+ T cells specific for respiratory viruses
in the human lung. J Exp Med 2005; 202:
1433-42. Abstract: http://amedeo.com/lit.php?id=16301748
- Flint SJ, Enquist LW, Racaniello
VR, Skalka AM. Principles of virology. Molecular
biology, pathogenesis, and control of animal
viruses. 2nd Edition, ASM Press,
Washington, DC, USA, 2004
- Gamblin SJ, Haire LF, Russell
RJ, et al. The structure and receptor binding
properties of the 1918 influenza hemagglutinin.
Science 2004; 303: 1838-42. Epub 2004 Feb
5. Abstract: http://amedeo.com/lit.php?id=14764886
- Garcia-Sastre A, Egorov A,
Matassov D, et al. Influenza A virus lacking
the NS1 gene replicates in interferon-deficient
systems. Virology 1998; 252: 324-30. Abstract:
http://amedeo.com/lit.php?id=9878611
- Garcia-Sastre A. Antiviral
response in pandemic influenza viruses.
Emerg Infect Dis 2006 (in press).
- Goto H, Kawaoka Y. A novel
mechanism for the acquisition of virulence
by a human influenza A virus. Proc Natl
Acad Sci U S A 1998; 95: 10224-8. Abstract:
http://amedeo.com/lit.php?id=9707628
- Guillot L, Le Goffic R,
Bloch S, et al. Involvement of toll-like
receptor 3 in the immune response of lung
epithelial cells to double-stranded RNA
and influenza A virus. J Biol Chem 2005;
280: 5571-80. Epub 2004 Dec 3. Abstract:
http://amedeo.com/lit.php?id=15579900
- Hall WJ, Douglas RG Jr,
Hyde RW, Roth FK, Cross AS, Speers DM. Pulmonary
mechanics after uncomplicated influenza
A infection. Am Rev Respir Dis 1976; 113:
141-8. Abstract: http://amedeo.com/lit.php?id=1247227
- Hayden FG, Fritz R, Lobo
MC, Alvord W, Strober W, Straus SE. Local
and systemic cytokine responses during experimental
human influenza A virus infection. Relation
to symptom formation and host defense. J
Clin Invest 1998; 101: 643-9. Abstract:
http://amedeo.com/lit.php?id=9449698
- Heath WR, Belz GT, Behrens
GM, et al. Cross-presentation, dendritic
cell subsets, and the generation of immunity
to cellular antigens. Immunol Rev 2004;
199: 9-26. Abstract: http://amedeo.com/lit.php?id=15233723
- Hemmes JH, Winkler KC, Kool
SM. Virus survival as a seasonal factor
in influenza and polimyelitis. Nature 1960;
188: 430-1. http://amedeo.com/lit.php?id=13713229
- Horner GJ, Gray FD Jr. Effect
of uncomplicated, presumptive influenza
on the diffusing capacity of the lung. Am
Rev Respir Dis 1973; 108: 866-9. http://amedeo.com/lit.php?id=4741881
- Hulse DJ, Webster RG, Russell
RJ, Perez DR. Molecular determinants within
the surface proteins involved in the pathogenicity
of H5N1 influenza viruses in chickens. J
Virol 2004; 78: 9954-64. Abstract: http://amedeo.com/lit.php?id=15331729
- Ishikawa E, Nakazawa M, Yoshinari
M, Minami M. Role of tumor necrosis factor-related
apoptosis-inducing ligand in immune response
to influenza virus infection in mice. J
Virol 2005; 79: 7658-63. Abstract: http://amedeo.com/lit.php?id=15919918
- Johansson BE, Bucher DJ,
Kilbourne ED. Purified influenza virus hemagglutinin
and neuraminidase are equivalent in stimulation
of antibody response but induce contrasting
types of immunity to infection. J Virol
1989; 63: 1239-46. Abstract: http://amedeo.com/lit.php?id=2915381
- Katz JM, Lim W, Bridges CB,
et al. Antibody response in individuals
infected with avian influenza A (H5N1) viruses
and detection of anti-H5 antibody among
household and social contacts. J Infect
Dis 1999; 180: 1763-70. http://amedeo.com/lit.php?id=10558929
- Katze MG, DeCorato D, Krug
RM. Cellular mRNA translation is blocked
at both initiation and elongation after
infection by influenza virus or adenovirus.
J Virol 1986; 60: 1027-39. Abstract: http://amedeo.com/lit.php?id=3023655
- Kondo S, Abe K. The effects
of influenza virus infection on FEV1 in
asthmatic children. The time-course study.
Chest 1991; 100: 1235-8. Abstract: http://amedeo.com/lit.php?id=1935277
- Lawrence CW, Braciale TJ.
Activation, differentiation, and migration
of naive virus-specific CD8+ T cells during
pulmonary influenza virus infection. J Immunol
2004; 173: 1209-18. Abstract: http://amedeo.com/lit.php?id=15240712
- Lawrence CW, Ream RM, Braciale
TJ. Frequency, specificity, and sites of
expansion of CD8+ T cells during primary
pulmonary influenza virus infection. J Immunol
2005; 174: 5332-40. Abstract: http://amedeo.com/lit.php?id=15843530
- Legge KL, Braciale TJ. Accelerated
migration of respiratory dendritic cells
to the regional lymph nodes is limited to
the early phase of pulmonary infection.
Immunity 2003; 18: 265-77. Abstract: http://amedeo.com/lit.php?id=12594953
- Levandowski RA, Gerrity
TR, Garrard CS. Modifications of lung clearance
mechanisms by acute influenza A infection.
J Lab Clin Med 1985; 106: 428-32. Abstract:
http://amedeo.com/lit.php?id=4045299
- Lipatov AS, Andreansky S,
Webby RJ, et al. Pathogenesis of Hong Kong
H5N1 influenza virus NS gene reassortants
in mice: the role of cytokines and B- and
T-cell responses. J Gen Virol 2005; 86:
1121-30. Abstract: http://amedeo.com/lit.php?id=15784906
- Little JW, Douglas RG Jr,
Hall WJ, Roth FK. Attenuated influenza produced
by experimental intranasal inoculation.
J Med Virol 1979; 3: 177-88. Abstract: http://amedeo.com/lit.php?id=479857
- Little JW, Hall WJ, Douglas
RG Jr, Mudholkar GS, Speers DM, Patel K.
Airway hyperreactivity and peripheral airway
dysfunction in influenza A infection. Am
Rev Respir Dis 1978; 118: 295-303. Abstract:
http://amedeo.com/lit.php?id=358877
- Mandelboim O, Lieberman
N, Lev M, et al. Recognition of haemagglutinins
on virus-infected cells by NKp46 activates
lysis by human NK cells. Nature 2001; 409:
1055-60. Abstract: http://amedeo.com/lit.php?id=11234016
- Matrosovich M, Tuzikov A,
Bovin N, et al. Early alterations of the
receptor-binding properties of H1, H2, and
H3 avian influenza virus hemagglutinins
after their introduction into mammals. J
Virol 2000; 74: 8502-12. Abstract: http://amedeo.com/lit.php?id=10954551
- Matrosovich MN, Matrosovich
TY, Gray T, Roberts NA, Klenk HD. Human
and avian influenza viruses target different
cell types in cultures of human airway epithelium.
Proc Natl Acad Sci U S A 2004; 101: 4620-4.
Abstract: http://amedeo.com/lit.php?id=15070767
- McCullers JA, Rehg JE. Lethal
synergism between influenza virus and Streptococcus
pneumoniae: characterization of a mouse
model and the role of platelet-activating
factor receptor. J Inf Dis 2002; 186: 341-50.
Abstract: http://amedeo.com/lit.php?id=12134230
- Mori I, Komatsu T, Takeuchi
K, Nakakuki K, Sudo M, Kimura Y. In vivo
induction of apoptosis by influenza virus.
J Gen Virol 1995; 76: 2869-73. Abstract:
http://amedeo.com/lit.php?id=7595397
- Olofsson S, Kumlin U, Dimock
K, Arnberg N. Avian influenza and sialic
acid receptors: more than meets the eye?
Lancet Infect Dis 2005; 5: 184-8. Abstract:
http://amedeo.com/lit.php?id=15766653
- Palladino G, Mozdzanowska
K, Washko G, Gerhard W. Virus-neutralizing
antibodies of immunoglobulin G (IgG) but
not of IgM or IgA isotypes can cure influenza
virus pneumonia in SCID mice. J Virol 1995;
69: 2075-81. Abstract: http://amedeo.com/lit.php?id=7884853
- Park CH, Ishinaka M, Takada
A, The invasion routes of neurovirulent
A/Hong Kong/483/97 (H5N1) influenza virus
into the central nervous system after respiratory
infection in mice. Arch Virol 2002; 147:
1425-36. Abstract: http://amedeo.com/lit.php?id=12111416
- Peiris JS, Yu WC, Leung
CW, et al. Re-emergence of fatal human influenza
A subtype H5N1 disease. Lancet 2004; 363:
617-9. http://amedeo.com/lit.php?id=14987888
- Piqueras B, Connolly J, Freitas
H, Palucka AK, Banchereau J. Upon viral
exposure myeloid and plasmacytoid dendritic
cells produce three waves of distinct chemokines
to recruit immune effectors. Blood 2005;
Abstract: http://amedeo.com/lit.php?id=16317096
- Renegar KB, Small PA Jr,
Boykins LG, Wright PF. Role of IgA versus
IgG in the control of influenza viral infection
in the murine respiratory tract. J Immunol
2004; 173: 1978-86. Abstract: http://amedeo.com/lit.php?id=15265932
- Sanz-Ezquerro JJ, Zurcher
T, de la Luna S, Ortin J, Nieto A. The amino-terminal
one-third of the influenza virus PA protein
is responsible for the induction of proteolysis.
J Virol 1996; 70: 1905-11. Abstract: http://amedeo.com/lit.php?id=8627716
- Schmitz N, Kurrer M, Bachmann
MF, Kopf M. Interleukin-1 is responsible
for acute lung immunopathology but increases
survival of respiratory influenza virus
infection. J Virol 2005; 79: 6441-8. Abstract:
http://amedeo.com/lit.php?id=15858027
- Shortman K, Liu YJ. Mouse
and human dendritic cell subtypes. Nat Rev
Immunol 2002; 2: 151-61. Abstract: http://amedeo.com/lit.php?id=11913066
- Silver ML, Guo HC, Strominger
JL, Wiley DC. Atomic structure of a human
MHC molecule presenting an influenza virus
peptide. Nature 1992; 360: 367-9. Abstract:
http://amedeo.com/lit.php?id=1448154
- Taubenberger JK. Influenza
virus hemagglutinin cleavage into HA1, HA2:
no laughing matter. Proc Natl Acad Sci U
S A 1998; 95: 9713-5. http://amedeo.com/lit.php?id=9707539
- To KF, Chan PK, Chan KF,
et al. Pathology of fatal human infection
associated with avian influenza A H5N1 virus.
J Med Virol 2001; 63: 242-6. Abstract: http://amedeo.com/lit.php?id=11170064
- Tran TH, Nguyen TL, Nguyen
TD, et al. Avian influenza A (H5N1) in 10
patients in Vietnam. N Engl J Med 2004;
350: 1179-88. http://amedeo.com/lit.php?id=14985470
- Treanor JJ. Influenza virus.
In: Mandell GL, Bennett JE, Dolin R, eds.
Mandell, Douglas, and Bennett's Principles
and Practice of Infectious Diseases. 6th
ed. Churchill Livingstone; 2004: 2060-2085.
- Tsitoura DC, Kim S, Dabbagh
K, Berry G, Lewis DB, Umetsu DT. Respiratory
infection with influenza A virus interferes
with the induction of tolerance to aeroallergens.
J Immunol 2000; 165: 3484-91. Abstract:
http://amedeo.com/lit.php?id=10975869
- Tumpey TM, Basler CF, Aguilar
PV, et al. Characterization of the reconstructed
1918 Spanish influenza pandemic virus. Science
2005; 310: 77-80. Abstract: http://amedeo.com/lit.php?id=16210530
- Uiprasertkul M, Puthavathana
P, Sangsiriwut K, et al. Influenza A H5N1
replication sites in humans. Emerg Infect
Dis 2005; 11: 1036-41. Abstract: http://amedeo.com/lit.php?id=16022777
- Ungchusak K, Auewarakul
P, Dowell SF, et al. Probable person-to-person
transmission of avian influenza A (H5N1).
N Engl J Med 2005; 352: 333-40. Abstract:
http://amedeo.com/lit.php?id=15668219
- Utell MJ, Aquilina AT, Hall
WJ, et al. Development of airway reactivity
to nitrates in subjects with influenza.
Am Rev Respir Dis 1980; 121: 233-41. http://amedeo.com/lit.php?id=7362132
- Weis W, Brown JH, Cusack
S, Paulson JC, Skehel JJ, Wiley DC. Structure
of the influenza virus haemagglutinin complexed
with its receptor, sialic acid. Nature 1988;
333: 426-31. Abstract: http://amedeo.com/lit.php?id=3374584
- Wiley JA, Cerwenka A, Harkema
JR, Dutton RW, Harmsen AG. Production of
interferon-gamma by influenza hemagglutinin-specific
CD8 effector T cells influences the development
of pulmonary immunopathology. Am J Pathol
2001a; 158: 119-30. Abstract: http://amedeo.com/lit.php?id=11141485
- Wiley JA, Hogan RJ, Woodland
DL, Harmsen AG. Antigen-specific CD8(+)
T cells persist in the upper respiratory
tract following influenza virus infection.
J Immunol 2001b; 167: 3293-9. Abstract:
http://amedeo.com/lit.php?id=11544317
- Wiley JA, Tighe MP, Harmsen
AG. Upper respiratory tract resistance to
influenza infection is not prevented by
the absence of either nasal-associated lymphoid
tissue or cervical lymph nodes. J Immunol
2005; 175: 3186-96. Abstract: http://amedeo.com/lit.php?id=16116209
- Wilson NS, Behrens GMN,
Lundie RJ, Systemic activation of dendritic
cells by TLR ligands or malaria infection
impairs cross cross-priming and anti-viral
immunity. Nat Immunol 2006 (in press)
- Woodland DL. Cell-mediated
immunity to respiratory virus infections.
Curr Opin Immunol 2003; 15: 430-5. Abstract:
http://amedeo.com/lit.php?id=12900275
- Woodland DL, Randall TD.
Anatomical features of anti-viral immunity
in the respiratory tract. Semin Immunol
2004; 16: 163-70. Abstract: http://amedeo.com/lit.php?id=15130500
- Woodland DL, Scott I. T
cell memory in the lung airways. Proc Am
Thorac Soc 2005; 2: 126-31. Abstract: http://amedeo.com/lit.php?id=16113480
- Yeldandi AV, Colby TV. Pathologic
features of lung biopsy specimens from influenza
pneumonia cases. Hum Pathol 1994; 25: 47-53.
Abstract: http://amedeo.com/lit.php?id=8314260
- Yen HL, Herlocher LM, Hoffmann
E, et al. Neuraminidase inhibitor-resistant
influenza viruses may differ substantially
in fitness and transmissibility. Antimicrob
Agents Chemother 2005; 49: 4075-84. Abstract:
http://amedeo.com/lit.php?id=16189083
- Yuen KY, Chan PK, Peiris
M, et al. Clinical features and rapid viral
diagnosis of human disease associated with
avian influenza A H5N1 virus. Lancet 1998;
351: 467-71. http://amedeo.com/lit.php?id=9482437
- Yuen KY, Wong SS. Human infection
by avian influenza A H5N1. Hong Kong Med
J 2005; 11: 189-99. Abstract: http://amedeo.com/lit.php?id=15951584
|
|
|
|
|
