|
第三章:流行性感冒病毒學
流行性感冒屬於正粘病毒科。正粘病毒包括:甲型、乙型及丙型流感病毒和霍高病毒Thogovirus
(在蜱的)。其中甲型及乙型流感病毒在人類間較為普遍。
甲型及乙型流感病毒主要的抗原體為血球凝集素(H或HA)和神經氨酸酵素 (N或NA)這兩種橫穿膜的糖蛋白。這兩種糖蛋白都有不同的抗原性,甲型流感病毒可根據十六種血球凝集素(H1至H16)及九種神經氨酸酵素(N1至N9)再作分類。每種流感病毒的正式命名包括:該病毒的類型(甲型或乙型),病毒的宿主(如宿主為人類則不需寫明),分離病毒的地方、編號、年份,以及H和N蛋白的類型。例如:A/goose/Guangdong/1/96
(H5N1)
流感病毒通常經飛沫傳播,接著感染呼吸管道內的黏膜,並穿透黏液層,進入呼吸道的表層細胞。流感病毒的繁殖速度很快,由感染細胞至散出細胞外只需六小時。流感病毒中的部份蛋白,如融合縮氨酸及NS2,能作為毒素以促進病毒的繁殖。病毒複製的初期或會引起細菌的快速繁殖(以肺炎鏈球菌,金黃葡萄球菌,及嗜血桿菌較為普遍)。(可參照「發病」一章)
結構
流感病毒為核糖核酸(RNA)病毒,由一個多樣性的外層包裹著,它的直徑約120nm。血球凝集素和神經氨酸酵素則覆蓋著病毒的表面。(Figure
1)
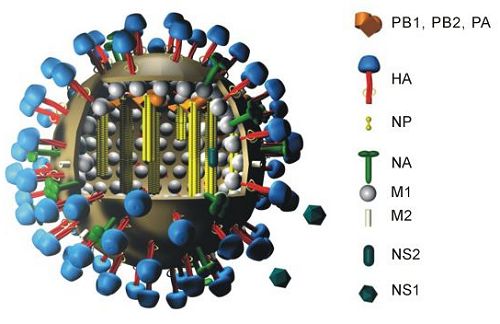
Figure
1. Structure of an influenza A virus. Image
copyright by Dr. Markus Eickmann, Institute
for Virology, Marburg, Germany. Used with permission.
. http://www.biografix.de
甲型及乙型流感病毒的基因組可分為八段,每段均被核殼蛋白質包裹著。它們合起來成為核糖核蛋白(RNP),而一段基因可被譯作一個有重要功能的蛋白:
- 聚合酵素B2蛋白 (PB2)
- 聚合酵素B1蛋白 (PB1)
- 聚合酵素A蛋白 (PA)
- 血球凝集素 (H或HA)
- 核殼蛋白 (NP)
- 神經氨酸酵素 (N或NA)
- 間質蛋白 (M):M1只在甲型流感病毒找到,它組成間質。M2作為離子管道,用以降低或保持內涵體的酸鹼度。
- 非結構性蛋白 (NS);NS2的功用為假定的蛋白
活性的RNA-RNA聚合酵素由PB2、PB1和PA形成,它們用作複製及抄寫 RNA。它與RNP連接著,具有核酸內切酵素的活性。NS1和NS2蛋白具有調整作用,用以促進病毒成份在被感染細胞內的形成。
病毒的表層為一雙層脂肪膜,來自生產病毒的細胞。膜上有著由HA,NA及M2蛋白組成的突出物。該脂肪膜包圍著由M1蛋白組成的間質。
丙型流感病毒的基因組只有七段,它的表層只有一種糖蛋白。因它對人類的致病性低,本書將不會對它作詳細討論。
血球凝集素
血球凝集素(HA或H)為糖蛋白,有兩或三個糖連結點。它的粒子重量約為76000。它在脂肪膜中伸延,因此該擁有最少五個抗原區域的主要部份能呈現在病毒表層的外側。HA用作一個感受器,它與唾液酸(N-acetylneuraminic
acid)結合,並引起膜融合使病毒內的粒子得以貫穿。血球凝集素為流感病毒主要的抗原體。其抗原點為A,B(帶有感受器結合點),C,D及E。抗原點在粒子的前端呈現,
而尾端則埋藏在脂肪膜層。HA粒子的軀幹包括莖部及遺傳因子融合地區。當病毒要感染一個新細胞時,該地區會用以引起膜融合。在低
pH值時,融合縮氨酸被移到內部。HA形成三聚物或,而數個三聚物或能形成一個融合細孔。
抗原點的突變會抑制抗體與抗原的結合,因此一種新類型的流感病毒能在未經免疫的人群間散播。這個現象稱為抗原飄變。造成抗原飄變的突變在分子層面上解釋了在冬天時於溫帶地區帶來季節性流感高峰期的現象。中和性抗體的生產帶來對HA抗原點的免疫性。抗體的生產是人類對抗感染的基礎。當一種新病毒在全球爆發時,有時候亦會帶來年長人士的部份交叉免疫。
抗原轉變──亦稱為基因組重新排列組合(ressortment)或只是重新排列組合──當HA在病毒間交換時發生,例如H5取代了H1,形式一個鑲嵌或嵌紋病毒。當同一個細胞被兩種流感病毒感染,而兩種病毒的基因段在複製時互相交換,就會形成抗原轉變。
這基因重新排列組合的現象在水鳥間甚為普遍,尤其在鴨的身上。雖然雀鳥受感染後甚少發病,但病毒能在雀鳥的糞便中生存數個月。
神經氨酸酵素
和HA相似,神經氨酸酵素(NA或N)為一種糖蛋白,亦同樣呈現在病毒表層的外側。它結合成為四聚物,粒子重量平均約為220,000。NA粒子的主要部份呈現在病毒表層的外側,在脂肪膜延展,並有個細小的細胞質的尾部。
NA用作一種酵素,將在細胞表面上的HA粒子、其他NA粒子、糖蛋白及糖脂類上的唾液酸切開。它亦作為一個重要的抗原點,並且似乎是令病毒穿越呼吸管道表層細胞上的黏液層的必要部份。
基因飄變亦會在NA上發生。NA帶有幾個重要的氨基酸,當它們發生突變,就能抵抗神經氨酸酵素抑制劑。已被發現的有關突變包括:
以上英文字母代表一種氨基酸(R,精氨酸;K,離氨酸;H,組氨酸;Y,酪氨酸;E,谷氨酸;V,纈氨酸):前面的英文子母代表原本的氨基酸,後面的代表突變後的氨基酸。
當在位於292的精氨酸(R)被離氨酸(K)取代,就會造成完全的抵抗性。該由R到K的突變源於N基因中的一個單核糖轉換:由AGA轉為AAA。該位置292的突變因此重要,因為它不但造成對奧斯他偉的抗藥性,亦會對扎那米韋及兩種新前趨藥物產生抵抗。
M2蛋白
當病毒分子在內涵體被收時,M2離子管道的活性會被提升,離子會湧入粒子內,降低pH值。因此,HA-M1的連結被干擾,令粒子打開,使HA內的融合縮氨酸被轉移,並且令HA與內涵體膜的內側融合。病毒的核糖蛋白被釋放到細胞間質,並且被運到細胞核內。在細胞核內,該複合物被干擾,發動病毒RNA的生產。
M2蛋白的活動會被金剛胺、金剛乙胺及有關物質抑制。
NS1的可能功用
人類的信使RNA在5'端帶有一個多A尾部。粒子重量約為26,000的NS1形式二聚物,抑制帶有多A尾部的信使RNA移出細胞核的輸送,籍以使病毒RNA優先被輸送到核糖體並且被翻譯。NS1亦有可能抑制前體信使RNA的剪接。此外,NS1很可能能夠壓制被感染細胞內的干擾反應,使得病毒的生產不被減弱。
NS2的可能功用
NS2為細小分子,分子重量約為11,000。在病毒內,它或許是與M1蛋白結合。它的功用相信是促進將新形成的RNA由細胞核輸送到間質,以加快病毒的生產。
複製週期
病毒吸附
流感病毒將HA的外端的頂部固定在細胞上的糖蛋白及糖脂類上的唾液酸上,以結合到細胞的表面。唾液酸與倒數第二個半乳糖的連結為α
2,3(雀鳥上)或α 2,6(在人類上),取決了宿主的專門性。因為帶有唾液酸的炭水化合物會在生物上的數種細胞上呈現,病毒的結合特性就能解釋到病毒會感染多種細胞的現象。
病毒的進入
當病毒附在細胞後,它就會經披網格蛋白衣受體介導內吞作用被吸收進細胞內。當病毒被帶進細胞內,披網格蛋白就會被釋放,那包裹著整個病毒的小囊會與內涵體融合。通常該舌噬體內的pH值會逐步降低,然後將小囊內的物體消化。
病毒脫殼
M2蛋白會在pH值到達一定水平後將其下降停止,使HA上的部份融合縮氨酸釋放出來。這樣HA就能夠與小囊的膜融合,使核糖蛋白(RNPs)經上述方法釋放到細胞間質。離子由內涵體湧入病毒,使得不同的病毒蛋白的連結切斷。M1蛋白的聚集被干擾,RNPs不再附在M1蛋白複合物上。病毒脫殼會在病毒附在細胞後的二十至三十分鐘內完結。
病毒RNA及病毒蛋白的合成
RNPs被輸送到細胞核內,聚合酵素複合物與RNA結合,以核酸內切酵素活動將病毒RNA劈開,並同時導致延長。病毒RNA的生產會被NP所限制支持mRNA的生產。它們兩者都會被輸送到細胞間質,病毒蛋白在核糖體內產生。部份病毒RNA被細胞內的酵素接合,因此最終病毒蛋白,如M1和NS2,能在不會再被劈開的情況下被合成。部份新合成的病毒蛋白被輸送到細胞核內,並與病毒RNA結合,形成RNPs。其餘新形成的病毒蛋白則在內質網及高基氏體被加工、醣化。這些被修改後的蛋白質被輸送到細胞膜及黏在雙層脂肪膜上。當它們在細胞膜上累積到一個足夠的濃度後,RNPs及M1蛋白會聚集及壓縮,以形成一個病毒粒子。最後,該病毒粒子會從細胞膜被擠出,經神經氨酸酵素活性釋放到細胞外。
由病毒的進入至新病毒粒子的形成,所需時間平均約六小時。
病毒脫落及傳染性
免疫組織學的圖片顯示,病毒生產細胞集中於在呼吸道上的黏膜層、腸藏,甚至在內皮細胞層、心肌及腦部。每一毫升的鼻分泌物能放出數以百萬計的病毒,因此一個0.1?l的懸浮粒子含有多於一百粒病毒。一個流感病毒的HID(使人受感染的劑量)約在100至1000個病毒粒子之間。至少在流感病毒感染的早期,病毒亦能在血液及其他體液內找到。
流感病毒傳染性的維持取決於溫度、pH值、水的鹽度及紫外線幅射。在攝氏4度時,在水中的流感病毒的傳染性的半衰期為2至3星期。因應病毒雙層脂肪膜的結構,病毒在一般環境因素下的生存能力會較短。
流感病毒的傳染力能輕易被酒精消毒劑、氯及乙醛消除。以現時所知,在攝氏70度的環境下,只需數秒就能破壞病毒的傳染力。
|
